Neurons are characterized by their remarkable ability to propagate signals rapidly over large distances. The primary mechanism by which they do this is called an action potential, an electrical pulse/spike that travel down nerve fibers.
Neurons are able to encode and transmit information and behavior by firing sequences of these spikes in various temporal patterns. This first chapter is dedicated to the understanding of neural coding, which will involve the measurement and characterization of how stimulus attributes(light, sound, etc / motor actions, arm movements, etc) are represented by these action potentials.
Most of this study takes the form of enacting some stimulus, and observing the response thereof. This is called Neural Encoding, where the stimulus is the input and the response is the output. An example of this would be to catalog how neurons respond to a wide variety of stimuli, and then construct models that attempt to predict responses to other stimuli.
Neural Decoding on the other hand refers to the opposite, given a response and attempting to understand or reconstruct the stimulus that caused it. However that is discussion for a later chapter.
This chapter will focus on the underlying biological principles on how neurons generate their response, and how such activity can be recorded.
Properties of Neurons
Neurons are essentially specialized cells that are capable of generating an electrical signal given a certain chemical input, or by receiving some other information from another input.
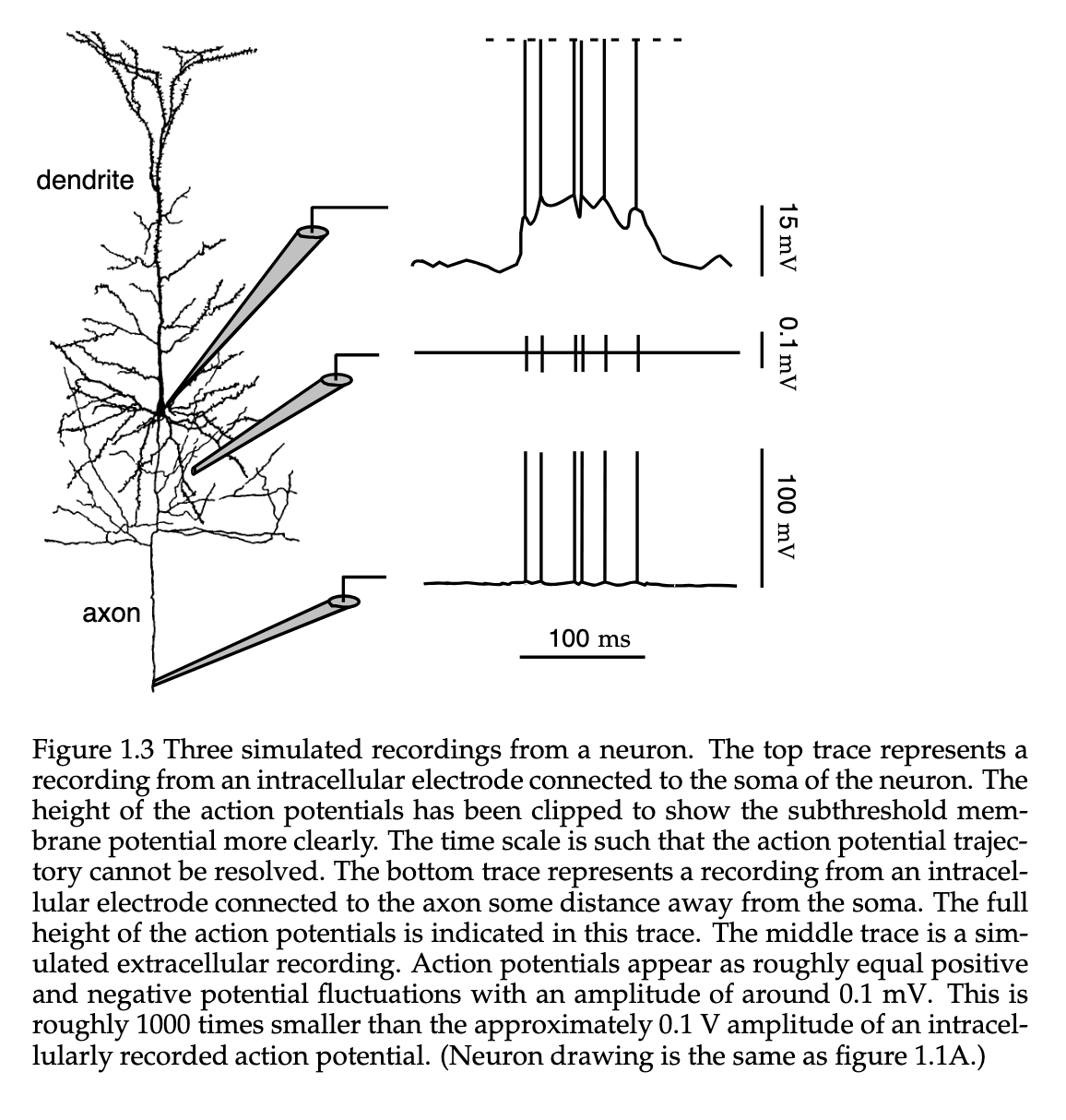
One of the more important specializations seen in the Neuron is that of the dendrite. These receive inputs from other neurons and the axon that carries the neuronal output to other cells. They are characterized by their elaborate “branching” structure gives them the ability to receive inputs from many other neurons through their synaptic connections. In figure 1.1, you can see that for A and C, the cortical pyramidial neurons have thousands of synaptic inputs. and in the special cell called a cerebellar Purkinje cell, it has over a hundread thousand( however this figure does not convey the full extent of the axon network ).
Axons can travel extreme distances from a single neuron to other areas in the brain, and in some cases the entire body. For a typical mouse, it is estimated that there is about 40mm of axon “cable” stemming from the cortical neurons, and about 4mm of total dendritic cable in their branched dendritic trees. An average axon has about 180 synaptic connections with other neurons per mm of length, while the dendritic trees have about 2 synaptic inputs per . The average cell soma has a range in diameter of about 10 to 50 .
Beyond the physical features of the neuronal components, there are associated physiological specializations. Namely, Neurons use a variety of membrane spanning Ion Channels that allow for ions ( mostly sodium , calcium, potassium , and chloride ) to move in and out of the cell. Put simply, these channels are like gates that open and close to allow ions to flow in and out of the cell. The movement of these ions across the membrane is what generates the electrical signals that neurons use to communicate with each other.
The relevant signal that dictates the behavior of the nervous system is the difference in these electrical potentials, specifically between the interior of a neuron and its surrounding extracellular medium. Under certain resting conditions, this potential is about relative to the surrounding bath, which is understood to be , in addition to the cell being polarized.
Ion Pumps at the cell membrane can maintain these concentration gradients, which support the Membrane Potential difference, as this would be a difficult state to maintain without.
A good example of this phenomena is the case of , which contains much higher concentrations outside of a neuron than inside it. In contrast, is found at a much higher rate inside of the neuron than outside in the extracellular medium. Because of these voltage and concentration gradients, Ions will consistently flow in and out of the cell, and this is what generates the electrical signals that neurons use to communicate with each other.
The current flowing out of the cell in the form of positively charged Ions(or the inverse, with negatively charged Ions flowing INTO the cell) goes through the open Ion Channels, making the Membrane Potential more negative. We call this process Hyperpolarization, and the opposite effect ( current flowing into cell changes membrane potential to a less negative and even positive value ) is called Depolarization.
When a given neuron is depolarized to a point where it raises the membrane potential above a certain threshold, it initiates a positive feedback process and leads to the neuron generating an action potential. This action potential is roughly a fluctuation across the cell membrane, lasting for about . These depend not only on the membrane potential, but also the recent history of that given neuron. For a very short period after a neuron has generated an action potential, is is near impossible for another spike to be generated. We call this the Absolute Refractory Period, which on average lasts some tens of milliseconds after a given spike.
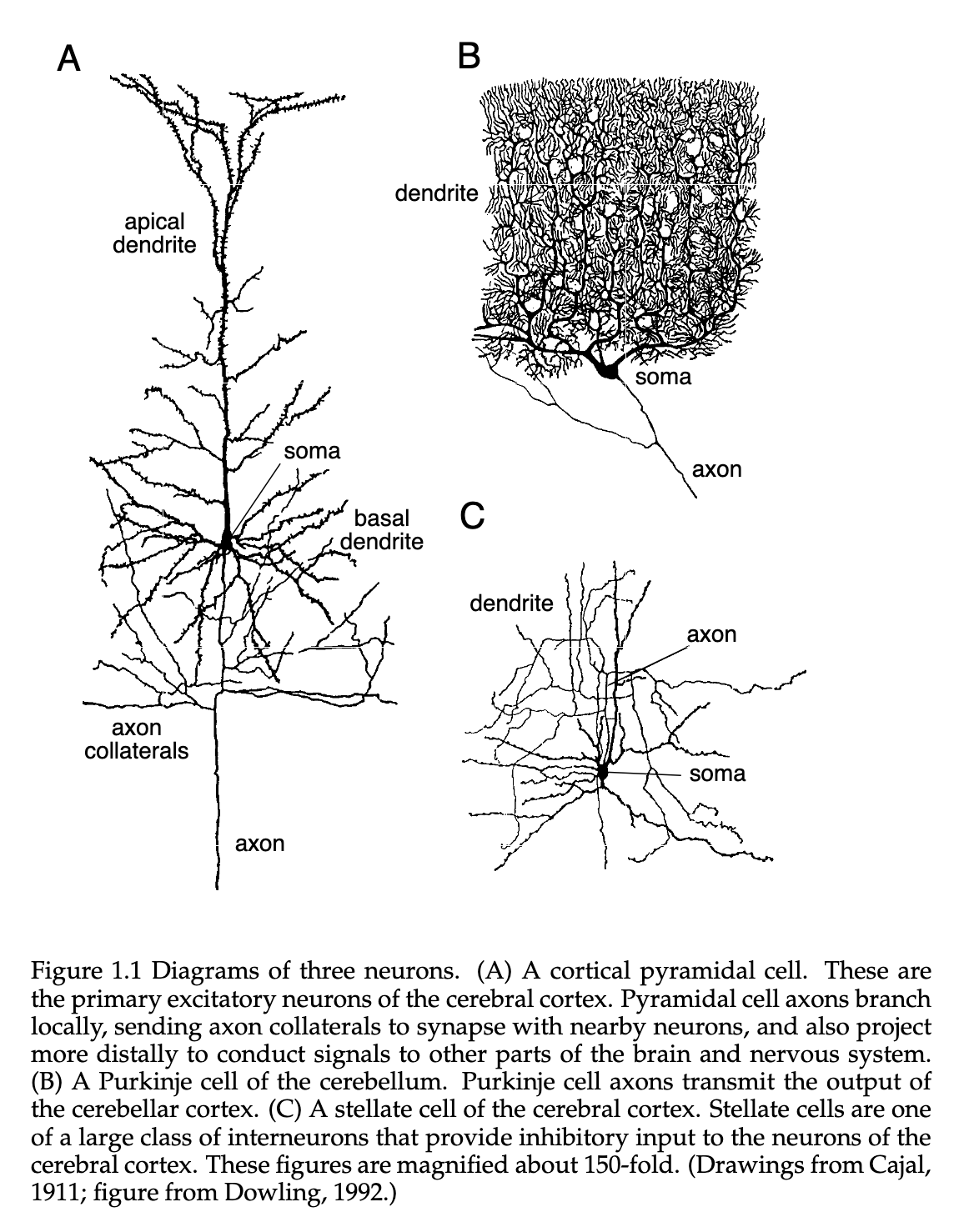
Action Potentials are extremely important to our understanding of the brain, as they are the only electrical fluctuation that can propagate over a vast distance. Other kinds of Subthreshold Membrane Potential fluctuations are very limited in their ability to travel beyond a distance of . Interestingly, Action Potentials are electrically regenerated as they pass along an axon process, similar to how a landline might “boost” the signal of a phone transmission after a certain distance. This mechanism allows them to travel rapidly over much larger distances without attenuation.
As discussed earlier above, Axons connect to dendrites through the use of Synapses, where the axon signal terminates at the synapse where the Voltage Transient of a Action Potential opens Ion Channels. This triggers a release of a large influx of , leading to the release of various Neurotransmitters( which you can see on figure 1.2B).
A neurotransmitter will then bind to a receptor on the postsynaptic(side that receives the signal) side of the synapse. This causes certain Ion Channels to open, which, depending on the nature of the ion flow, leads to the synapses having an excitatory/depolarizing/inhibitory/hyperpolarizing effect on the postsynaptic neuron.
How We Record Neuronal Responses
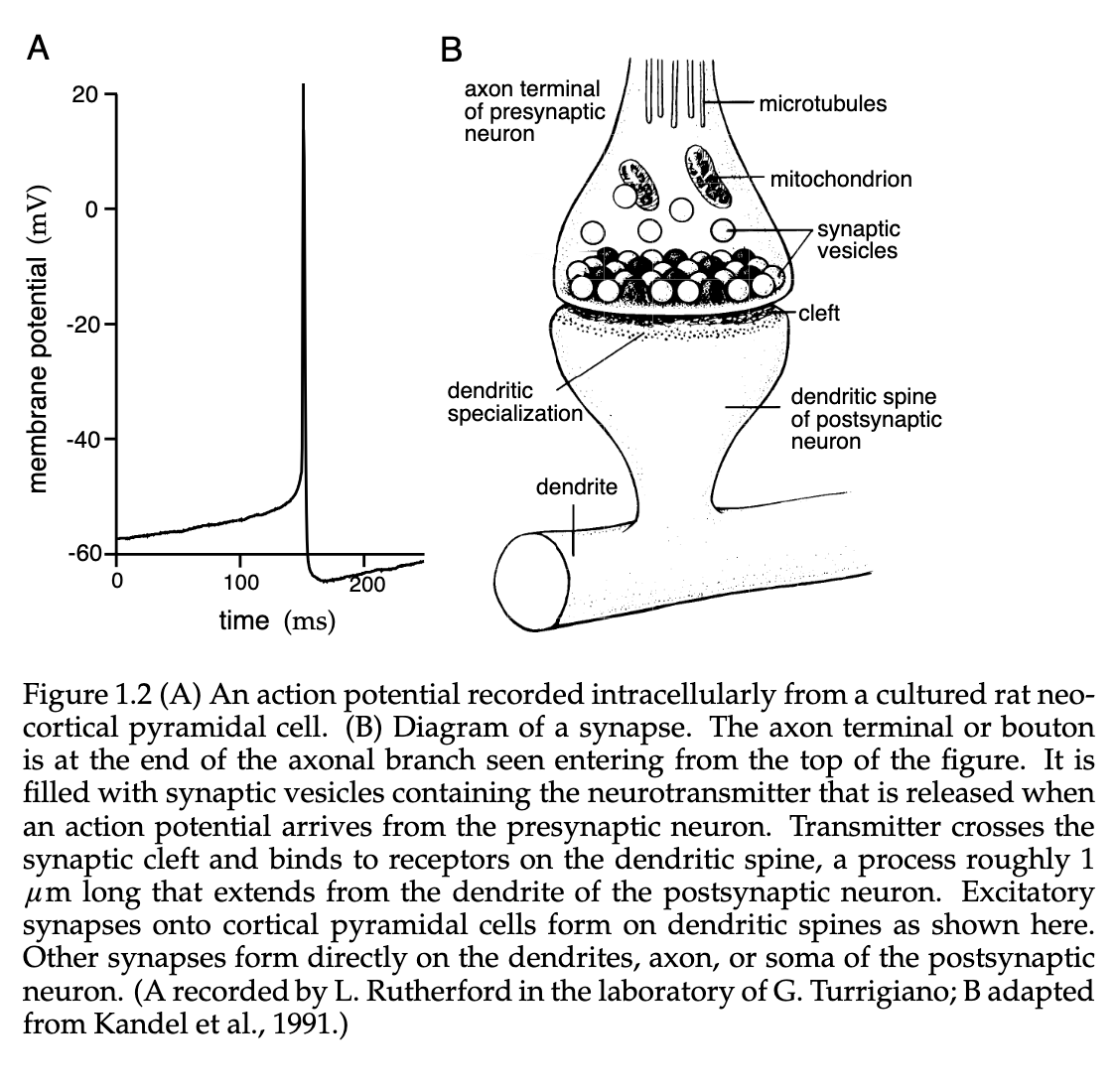
The figure below shows several different methods for using Electrodes to record neuronal information. The two primary methods to record this information are the intracellular and extracellular approaches, but this can also be done using an optical method.
The Membrane Potential can be recorded using the intracellular approach by insert a special hollow glass electrode filled with some conducting electrolyte to a given neuron, which then yields a reading by comparing the potential it records with that of a reference electrode place in the surrounding Extracellular Space.
We can record the axon potential by implementing a extracellular approach. This approach is typically used for in vivo experiments, such as behaving animals. The process for an extracellular approach involved placing an electrode near a Neuron, but not penetrating the cell membrane. A common case is for in vitro preparations, in such cases like dealing with slices of neural tissue.